
Accueil / Publications / Fiches bioagresseurs / Le SBR, le RTD et les cicadelles vectrices

Le Syndrome des basses richesses (causé par Candidatus Arsenophonus phytopathogenicus) et le Rubbery Taproot Disease (causé par Candidatus Phytoplasma solani) sont des maladies bactériennes transmises par les cicadelles de la famille des cixiidés. Ces deux pathogènes peuvent coexister dans une même plante, comme observé en Allemagne et sporadiquement en Bourgogne dans les années 2000. En raison de leurs symptômes foliaires proches et de leur biologie similaire, ces pathogènes sont parfois confondus. Ainsi, selon les sources, le terme "Syndrome des basses richesses" (SBR) peut désigner soit le complexe de ces deux agents pathogènes, soit spécifiquement la maladie causée par Candidatus Arsenophonus phytopathogenicus.
Dans la suite de cette fiche, "Syndrome des basses richesses (SBR)" désignera la maladie causée par C. Arsenophonus phytopathogenicus.
Dernière mise à jour octobre 2025
Bien qu’une partie de leur nom soit similaire, ces deux bactéries n'appartiennent pas au même groupe phylogénétique. Candidatus Arsenophonus phytopathogenicus appartient au phylum des Proteobacteria, tandis que Candidatus Phytoplasma solani appartient au phylum des Tenericutes. En effet, le terme "Candidatus" n'indique pas un groupe phylogénétique homogène, mais regroupe des bactéries qui ne peuvent pas être cultivées, rendant ainsi leur caractérisation plus complexe (Bertaccini et al. 2022).
Selon les pays, ces pathogènes sont émergents, ré-émergents ou en expansion. La recherche est donc très active sur ces maladies et les informations listées sur cette fiche résument l’état des connaissances en octobre 2025. Elles sont susceptibles d’évoluer avec l’avancée des recherches.
Le syndrome des basses richesses a été détecté pour la première fois sur betterave en Europe, en Bourgogne et en Franche-Comté en 1991. Il est associé à la protéobactérie Candidatus Arsenophonus phytopathogenicus (ARSEPH) qui a été identifiée au début des années 2000 (Bressan et al. 2012).
Cette bactérie est présente uniquement en Europe (EPPO 2025a). Elle est en forte expansion en Hongrie depuis 2005, en Allemagne depuis 2009 et en Suisse depuis 2017. En 2012, l'EPPO (European and Mediterranean Plant Protection Organization) déclare un risque phytosanitaire élevé pour les zones de culture de betterave dans toute l'Europe, recommandant la surveillance des cultures.
Cette maladie s'étend d’environ 15 km par an. La Suisse a connu une forte augmentation des surfaces en 2020 à la suite d’une crise en co-infection avec la jaunisse (Mahillon et al. 2022). En Allemagne, les surfaces concernées ont doublé entre 2019 et 2020 pour atteindre 40 000 ha touchés et la maladie se répand rapidement dans le sud et l’est, menaçant l’industrie de la betterave (Kais et al. 2023). Le SBR n’était plus détecté en France depuis la fermeture de la sucrerie d’Aiserey en 2007, jusqu’à sa réapparition en Alsace en 2023 sur des surfaces très limitées.
Les bactéries du genre Arsenophonus sont majoritairement des endosymbiontes d’arthropodes, mais certaines peuvent aussi agir comme pathogènes de plantes comme ARSEPH (Bressan 2014).
Ces bactéries peuvent être des symbiontes obligatoires, indispensables à la survie de l’insecte hôte, ou des symbiontes secondaires, comme c’est le cas pour ARSEPH) Des espèces voisines peuvent modifier les aptitudes biologiques ou le comportement de leurs hôtes. L’impact éventuel d’ARSEPH sur les cicadelles n’est pas connu à ce jour (Bressan 2014). Aucune anomalie des cellules infectées chez l’insecte n’a été observée ce qui suggère que la bactérie ne nuirait pas à son hôte (Bressan et al. 2009).
Les symptômes de SBR sur betteraves sont caractérisés par le jaunissement et la nécrose des feuilles les plus vieilles et des repousses de feuilles asymétriques, chlorotiques et lancéolées (Figure 1). Les symptômes sont diffus, sur la parcelle entière, on ne distingue généralement pas de rond ou de tâches. Des brunissements vasculaires sur la racine (Figure 2) sont observées, correspondant à une nécrose des tubes de phloème, une lignification de la paroi cellulaire et un dépôt de phénols (métabolites secondaires couramment impliqués dans les mécanismes de défense des plantes) dans le lumen (Gatineau et al. 2002). Les racines conservent en revanche une taille normale (Gatineau et al. 2002).

Figure 1 : à gauche des feuilles issues de plantes malades, à droite des feuilles saines.

Figure 2 : à gauche une betterave saine, à droite une betterave avec des traces de noircissement du phloème dues au SBR (ITB)
Les symptômes apparaissent 2 à 4 mois après l’infection des betteraves au champ (Sémétey et al. 2007) mais seulement 52 jours après l’inoculation en conditions contrôlées (Mahillon et al. 2022).
ARSEPH se trouve dans les cellules du phloème auquel il est limité, principalement dans les racines et les pétioles des vieilles feuilles. Elle n'est pas détectée par PCR dans le limbe des feuilles ni dans les pétioles des jeunes feuilles (Mahillon et al. 2022).
C. Arsenophonus phytopathogenicus entraine une diminution de la biomasse racinaire jusqu’à 25 % et de la richesse jusqu’à 5 % (Bressan et al. 2008). La virulence de la maladie varie selon les parcelles et les campagnes, mais presque toutes les plantes peuvent être concernées dans des parcelles lourdement touchées.
Le principal hôte connu est la betterave sucrière. En Allemagne et en Suisse, ARSEPH a récemment été détecté sur la pomme de terre, se manifestant par un retard de croissance ainsi qu’un jaunissement et un enroulement des feuilles, dans les zones où les betteraves sont fortement touchées (Behrmann et al. 2023 ; Therhaag et al. 2024 ; Mahillon et al. 2025). Des oignons symptomatiques et non symptomatiques se sont aussi déclarés être porteurs d’ARSEPH en Allemagne (Therhaag et al. 2024). Récemment, il a été montré que le vecteur du SBR peut réaliser une partie de son cycle et transmettre ARSEPH à la carotte Daucus carota en Allemagne sur lesquelles ont été constatés un jaunissement ou rougissement des feuilles (Witczak et al. 2025).
C. Phytoplasma solani (PHYPSO) est également transmis à la betterave par des cixiidés. Il peut être présent seul ou en co-infection avec C. Arsenophonus phytopathogenicus. C’est un phytoplasme, c’est-à-dire une bactérie sans paroi. C’est un parasite obligatoire, qui ne peut pas se reproduire sans son hôte, qui se développe dans les cellules du phloème des plantes auquel il est limité ou dans les cellules d’insectes.
Il est notamment associé à la maladie nommée ‘Rubbery Taproot Disease’ (RTD) sur betterave et au stolbur sur pomme de terre. Cette maladie a été observée pour la première fois en Serbie dans les années 1960. En 2018, la Serbie a connu une importante épidémie de RTD. Les formes agressives de ce pathogène n’ont pas été détectées en France jusqu’à présent. En revanche, elles sont présentes à l’est du continent (Est de l’Allemagne, Serbie, Roumanie, Hongrie, Croatie, Autriche, Slovaquie).
Les symptômes du Rubbery Taproot Disease apparaissent autour de la deuxième quinzaine de juillet, environ un mois après l’infection au phytoplasme. Ils se caractérisent par une perte de turgescence suivie d’un jaunissement puis d’une nécrose des feuilles anciennes en commençant par leur bord. Progressivement, toutes les feuilles nécrosent. La racine est caoutchouteuse, mais ne comporte pas de traces foncées contrairement au SBR (Figures 3 et 4).

Figure 3 : Symptômes du Rubbery Taproot Disease (Ćurčić et al. 2021)

Figure 4 : une racine "molle" touchée par le RTD
L’intensité des symptômes causés par Candidatus Phytoplasma solani sur la betterave est très variable selon la souche présente et les conditions climatiques :
Par rapport à ARSEPH, ce pathogène entraîne une plus grande diminution de la biomasse racinaire des betteraves, de la richesse et une plus grande prolifération de feuilles (Bressan et al. 2007 ; Bressan et al. 2008). Ces pertes varient considérablement en fonction de la souche concernée. De plus, les betteraves caoutchouteuses peuvent être difficiles à couper, ce qui empêche l’extraction du sucre, rendant les betteraves non marchandes dans le pire des cas (Ćurčić et al. 2021).
C. Phytoplasma solani peut permettre à des pathogènes opportunistes de s’installer, notamment le champignon Macrophomina phaseolina, entrainant des pourrissements, alors qu’il est habituellement pathogène du soja, du sorgho ou de l’arachide. Dans le cas d’étés chauds et de sécheresses intenses, les pertes peuvent aller jusqu’à 100 % (Duduk et al. 2023). En Serbie, il a été constaté que le champignon est présent uniquement dans les champs de betteraves infectés de RTD. En conditions contrôlées, même en conditions optimales, le champignon n'infecte pas les betteraves si elles ne sont pas infectées par PHYPSO avant (Duduk et al., 2023).
PHYPSO est responsable de maladies sur un large nombre de plantes cultivées (stolbur sur tomate ou pomme de terre, maladie du bois noir sur vigne, maïs…), notamment les solanacées et les apiacées mais est également détectée sur des plantes spontanées (ortie, liseron, trèfle…). En tout, plus de 100 plantes hôtes ont été identifiées (EPPO 2025b). Mais, tout comme l’intensité des symptômes, la gamme d’hôtes dépend de la souche étudiée.
Différentes populations de cixiidés ayant des cycles et notamment des hôtes différents sont porteuses de souches de PHYPSO différentes (Langer et Maixner 2004). Les cicadelles provenant du liseron (Convolvulus arvensis) sont vectrices du RTD sur betterave tandis que celles provenant de l'ortie (Urtica dioica) sont responsables du stolbur sur pomme de terre (Bressan et al. 2007). Ces deux maladies sont associées à des souches différentes de PHYPSO distinguées par le séquençage de leurs gènes de ménages tuf codant le facteur d'élongation Tu, une protéine essentielle à la synthèse des protéines. Les souches de PHYPSO respectivement associées au liseron et à l'ortie (et donc au RTD sur betterave et au stolbur sur pomme de terre) sont tuf-b1 ou tuf-d et tuf-b2 ou tuf-a (Langer et Maixner 2004 ; Kosovac et al. 2023 ; Contaldo et al. 2021 ; Curcic et al. 2021). Au début des années 2000, c'est la souche tuf-b associée au stolbur de la pomme de terre qui est la plus répandue et abondante en Allemagne (Langer et Maixner 2004).
Il est donc nécessaire d’identifier le phytoplasme présent à l’échelle du sous-groupe ou de la souche pour l’épidémiologie. Néanmoins, la caractérisation moléculaire de cet agent pathogène reste complexe et doit se faire sur trois zones distinctes du génome (Eini et Varrelmann 2023).
Les cicadelles vectrices de ARSEPH et PHYPSO sont de la famille des Cixiidae de l'infraordre Fulgoromorpha. Comme les pucerons, elles appartiennent à l’ordre des Hémiptères : des insectes possédant un rostre et des ailes membraneuses. Environ 30 espèces de la famille des cixiidés sont présentes en Europe centrale. Les cixiidés sont des insectes piqueurs-suceurs dont les adultes se nourrissent de la sève élaborée dans le phloème des plantes sur les parties aériennes et les nymphes sur les racines puisqu'elles sont dans le sol.
De nombreuses espèces de cixiidés sont vectrices de virus et de bactéries d’importance économique majeure, comme la bactérie du bois noir sur la vigne ou du stolbur sur pomme de terre.
Pentastiridius leporinus (Figure 5) et Cixius wagneri sont porteuses de Candidatus Arsenophonus phytopathogenicus et ont été identifiées comme vecteurs du SBR en 2005 et 2006. P. leporinus a été désigné comme étant le vecteur économique d'ARSEPH sur betterave étant donné son abondance et son taux d'infection élevé pouvant aller jusqu’à 95 % (Sémétey et al., 2007 ; Bressan et al., 2009). En effet, cette espèce représentait plus de 97 % des cixiidés capturées en France par Bressan et al. (2007). De plus, les larves peuvent être observées facilement dans les champs symptomatiques en début d’automne (Figure 7).


Figure 5 : Cicadelles P. leporinus adultes sur betteraves (ITB).
La cicadelle Hyalesthes obsoletus (Figure 6) est porteuse du phytoplasme C. Phytoplasma solani responsable du RTD. P. leporinus est également capable de transmettre le phytoplasme (Behrmann et al. 2023 ; Therhaag et al. 2024). Néanmoins, cet agent pathogène n’a pas été détecté dans les P. leporinus capturés en Bourgogne en 2005 et 2006 (Bressan et al. 2007) mais uniquement dans les cicadelles H. obsoletus.

Figure 6 : Une cicadelle Hyalesthes obsoletus adulte, vectrice du RTD et du stolbur (KWS).
Récemment, la cicadelle Reptalus quinquecostatus a été identifiée comme vecteur du SBR et du RTD en Serbie (Duduk et al. 2023 ; Kreitzer et al. 2025). Dans ces zones, des taux d’infection de R. quinquecostatus à PHYPSO ont été mesurés à 67 % avec des infections aux souches tuf-b et tuf-d (Kosovac et al. 2023).
Reptalus cuspidatus a aussi été identifiée comme vecteur du RTD et du stolbur en Serbie, mais ayant a priori un rôle plus secondaire que R. quinquecostatus dans les épidémies majeures (Kosovac et al. 2023).
Plus récemment, Reptalus quinquecostatus, principalement présente en Europe centrale, a aussi été identifié comme vecteur d’ARSEPH donc du SBR sur betterave (Kreitzer at al. 2025).
Ces cixiidés ont longtemps été considérées comme des espèces univoltines, c'est-à-dire produisant une génération par an. Cependant, en 2022 en Allemagne, dans des conditions d'un été chaud et sec, un second vol de P. leporinus a été observé fin août, à la suite d'un premier vol début juillet puis d'un déclin (Berhmann et al. 2023). L'apparition de deuxièmes générations génère probablement une augmentation de la population et du risque de transmission des pathogènes et sera favorisée à l'avenir avec l'augmentation des températures.
Les adultes de Pentastiridius leporinus migrent sur la betterave sucrière en fin de printemps ou au début de l’été (Behrmann et al. 2023). En 2023, Behrmann et al. ont constaté que P. leporinus peut aussi compléter son cycle sur pomme de terre. Ainsi, on suppose que les adultes peuvent passer de la pomme de terre à la betterave et inversement de juin à septembre.
On observe des vols de mai jusqu'en août et parfois jusqu'en septembre si l'été est chaud et sec (Behrmann et al., 2022). Après l’accouplement, qui a lieu fin juin, les femelles pondent leurs œufs dans le sol à proximité des racines qui éclosent environ 2 semaines après (R. Pfitzer et al. 2022). Les femelles pondent en moyenne 139 œufs par individu sur betterave (Pfitzer et al. 2022). Le cycle de vie se décompose en cinq stades nymphaux et un stade adulte. Les nymphes se développent dans l’horizon supérieur du sol où elles se nourrissent sur les racines de betteraves jusqu’à l’arrachage. Dans les champs présentant des symptômes, on peut observer les larves blanches de P. leporinus en arrachant des racines de betterave. Elles sont facilement reconnaissables grâce au « plumeau » blanc visible à l’arrière et aux structures filamenteuses et cireuses blanches que les femelles et les larves produisent pour se protéger des prédateurs et réguler l'humidité. D'après Behrmann et al. (2022), ces filaments sont produits par les cinq stades nymphaux de P. leporinus ainsi que par les femelles.

Figure 7 : Une larve de P. leponirus sur une racine de betterave.
Ensuite, les stades larvaires plus âgés se nourrissent sur les racines de la culture suivante, généralement du blé d’hiver (Bressan, 2014). Les nymphes n'effectuent pas de diapause, elles restent actives à des basses températures en hiver, même à 0 °C. Leurs mouvements dans le sol sont en revanche limités par ces basses températures. Au-delà d'une température moyenne du sol de 8,6 °C, les nymphes se trouvent principalement dans la couche supérieure du sol (0-10 cm). En dessous de 5,6 °C, leur abondance augmente dans les couches inférieures (10-20 et 20-30 cm). Les cicadelles remontent la surface pour émerger l’été suivant et migrent sur betterave sucrière (Figure 6).
Une fois infectée, P. leporinus est capable de transmettre la bactérie responsable du SBR après quelques jours et reste infectieuse toute sa vie, d’une durée d’environ 30 jours (Bressan at al. 2009).
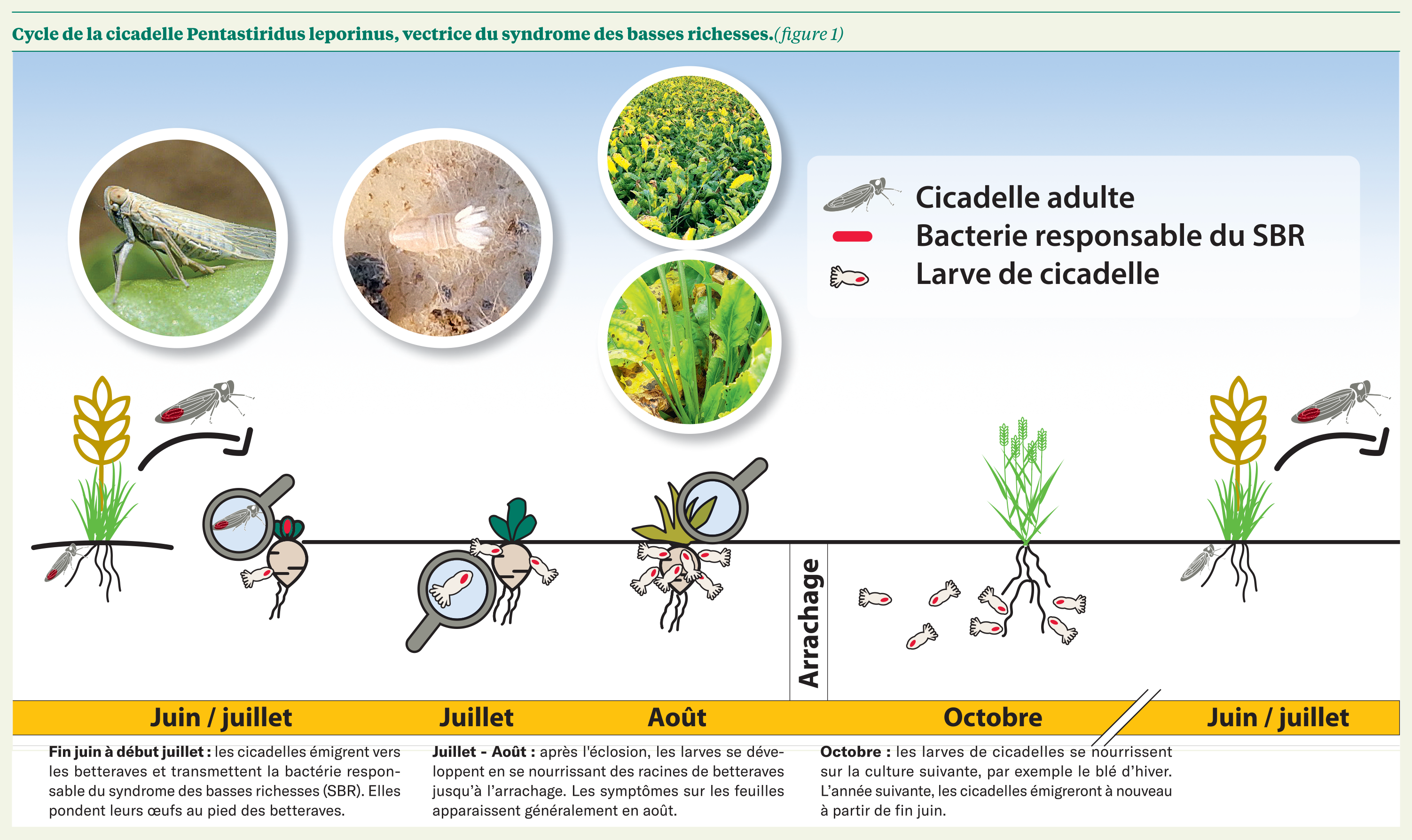
Figure 8 : Cycle de vie de Pentastiridus leporinus (Le Betteravier Français).
ARSEPH et PHYPSO sont exclusivement transmis par des insectes vecteurs. Sur betterave, ARSEPH est transmis par les cixiidés P. leporinus (principal vecteur), C. wagneri et R. quinquecostatus (en Europe centrale) (Sémétey et al. 2007). PHYPSO est transmis par H. obsoletus (principal vecteur), P. leporinus, R. quinquecostatus et R. panzeri (en Europe centrale).
La transmission d’ARSEPH, responsable du SBR, se fait à la fois horizontalement (via les plantes) et verticalement (de la femelle à sa progéniture) tandis que la transmission de PHYPSO, responsable du RTD, se fait uniquement horizontalement (via les plantes). Pour ce dernier, la transmission est moins efficace par P. leporinus que par H. obsoletus, ce qui fait de ce dernier le vecteur principal (Bressan et al. 2008).
La transmission horizontale d'ARSEPH et de PHYPSO est possible par les nymphes et les adultes de P. leporinus. Tout d'abord, l'insecte acquiert le pathogène par ses pièces buccales et s'infecte en exploitant la sève du phloème des plantes. Ensuite, le pathogène se propage dans le corps de l'insecte. ARSEPH est abondant dans la plupart des organes internes de P. leporinus avec la plus grande concentration dans les glandes salivaires (surtout chez les femelles) et les gonades des adultes (Bressan et al. 2009). Après cette période de latence, les insectes peuvent transmettre le pathogène à la plante hôte. P. leporinus reste infectée par ARSEPH toute sa vie, d'environ 30 jours (Bressan et al. 2009). Dans des conditions contrôlées, pour des cicadelles élevées en groupe, le taux d'infection des cicadelles P. leporinus par C. Arsenophonus phytopathogenicus reste constant, autour de 80 % (R. Pfitzer et al. 2022). Les adultes femelles vivent plus longtemps et sont de meilleurs vecteurs que les mâles, respectivement 64 % et 34 % d'efficacité de transmission, car la bactérie est plus abondante dans les glandes salivaires de femelles que des mâles (Bressan et al. 2008).
La transmission verticale d’ARSEPH a une efficacité assez faible de 30 % et joue donc un rôle secondaire dans le maintien de l'infection des populations de P. leporinus par rapport à la transmission horizontale (Bressan et al. 2009). Le processus de transmission verticale n'est pas connu à ce jour, il n'y a pas de preuve évidente d'infection des œufs en formation dans les ovaires (Bressan et al. 2012). La haute concentration d'ARSEPH dans les oviductes suggère que le pathogène entre dans l'œuf pendant sa transition dans l'oviducte ou que la bactérie adhère à la surface de l'œuf et entre dans l'insecte seulement après éclosion (Bressan et al. 2012).
H. obsoletus (Figure 7) est une cicadelle très polyphage. Les adultes sont retrouvés sur de nombreuses espèces, cultivées ou non. En revanche, il est moins évident de connaitre les espèces sur lesquelles les larves sont capables de faire leur développement complet (Kosovac et al. 2018). L’ortie et le liseron sont considérés comme les plantes hôtes majeures pour les adultes et les larves, dans l’agrosystème viticole. Les larves sont aussi capables de compléter leur cycle sur lavande et une partie de leur cycle sur le céleri, le maïs ou encore la sauge.
C. wagneri est également vectrice de Candidatus Phlomobacter fragariae, responsable de la chlorose marginale du fraisier. Elle a donc principalement été étudiée dans le contexte de la production de fraises jusqu’à présent. Cette cicadelle est capable de faire son cycle complet en tunnels de fraisiers et de se développer sur les renouées qui peuvent infester les parcelles de betteraves. Cette préférence d'hôte pourrait expliquer la plus grande fréquence de C. wagneri porteuses d'ARSEPH à l'automne qu'au printemps. Elles se nourriraient seulement dans un second temps sur les betteraves infectées par P. leporinus précédemment.
L'identification des cixiidés se fait par observation de la morphologie externe (tête, pâtes, ailes, structures génitales) ou par dissection. Par exemple, cette famille d'hémiptères a la particularité d'avoir des ailes disposées en toit au repos. Pour distinguer les espèces au sein des cixiidés, il existe de nombreux critères morphologiques : le nombre de carènes du mésotonum, le nombre d'épines sur le premier segment du tarse postérieur, les caractéristiques du scutellum, du vertex et du pronotum. Les cinq stades nymphaux (N1 à N5) sont identifiables par la largeur de la tête (capsule céphalique) (Pfitzer et al. 2022). En revanche, cette méthode nécessite une expertise et des insectes intacts pour déterminer l'espèce sur ces critères. P. leporinus a parfois été confondue avec P. beieri, du fait de leurs similitudes morphologiques (Gatineau et al. 2002).
Autrement, la méthode moléculaire de barcoding ADN permet d'identifier les espèces à partir de la séquence du gène mitochondrial cytochrome-oxydase I COI. Il est souvent nécessaire d'utiliser la méthode moléculaire pour différencier les mâles des femelles, notamment au stade d'œuf et de nymphe où il n'y a pas de différence morphologique entre les mâles et les femelles (Behrmann et al. 2022).
Identifier les espèces de cixiidés est nécessaire dans le cadre du monitoring puisque les espèces sont associées à des cycles de vie, des pathogènes et des maladies différentes. D'après Pfitzer et al. (2022), il est possible d'identifier les cixiidés capturées à l'aide de pièges collants jusqu'à 18 mois après conservation.
L'étude des cixiidés passe par des tests de choix de plantes, des tests de transmission de bactéries, de mortalité, de prédation, des élevages, du monitoring, de l'identification ou encore du séquençage. Pour la première fois en 2022, Pfitzer et al. a réussi à maintenir un élevage en masse de P. leporinus. Cinq générations ont été obtenues en deux ans avec plus de 20 000 individus élevés de l'œuf à l'adulte uniquement sur betterave sucrière. Cette avancée permet d’apporter de nombreuses informations sur la biologie de l’insecte.
A ce jour, il n'existe pas de méthode efficace de lutte contre les cixiidés vectrices du SBR et du RTD mais la recherche est très active sur le sujet dans toute l’Europe. Parmi les méthodes explorées dans le but de réduire les populations ou limiter l’impact de la maladie, on retrouve entre autres l'application de produits, le travail du sol, la rotation des cultures, le recours à des prédateurs naturels ou des variétés résistantes.
Concernant P. leporinus, vecteur principal du SBR :
Les recherches sur H. obsoletus, vecteur principal du RTD :
Les moyens de lutte visant à réduire l’impact des maladies :
En Allemagne, où le SBR et le RTD sont un problème majeur, une task force a été constituée en 2023 pour intensifier les travaux de recherche de solutions pour les agriculteurs.
Les efforts et les connaissances sont mutualisés à travers l’Europe par le biais de séminaires, de visites d’essais et autres échanges réguliers.
En France, le projet IPSEELON (Insectes Piqueurs-Suceurs Epidémiosurveillance et Lutte contre les Organismes Nuisibles), démarré en 2026, réunit notamment les filières pomme de terre (plants et de consommation) et betterave sucrière. Ce projet permettra d’apporter des connaissances sur la présence et le développement des cixiidés en France, ainsi que sur de potentielles méthodes de luttes (tolérance variétale, produits de biocontrôle, prédation).
Références
|

![]()
Agrément conseil de l’ITB à l’utilisation des produits phytosanitaires n° 7500002.
Assurance RC n° 05421646Y/1025.
Le portail EcophytoPIC recense les techniques alternatives à l’utilisation des produits phytopharmaceutiques.
Organisme agréé Crédit d'impôt Recherche
L'Institut Technique de la Betterave est
membre du réseau Acta
Institut Technique Agricole Qualifié
par le Ministère de l’Agriculture et de l’Alimentation
