
Accueil / Publications / Fiches bioagresseurs / Les jaunisses virales et leurs pucerons vecteurs
Les jaunisses sont des maladies virales transmises par des pucerons vecteurs. Les hivers doux qui précèdent la betterave sont propices à la multiplication des pucerons et au maintien des réservoirs viraux. L’impact des jaunisses virales sur le rendement est variable selon le type de virus impliqué et la date d’inoculation. En 2020 (deuxième année sans néonicotinoïde en traitement de semences), toutes les régions de production ont été touchées par la jaunisse, ce qui a entraîné d’importantes pertes de rendement. Cette fiche fait l’état des connaissances sur la biologie du pathosystème betterave/puceron/virus et des mécanismes épidémiologiques régissant la dissémination virale au champ.
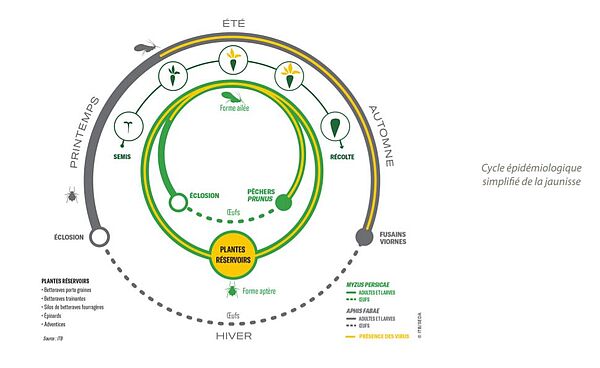
La jaunisse, telle qu’elle classiquement désignée dans le monde betteravier, est en réalité un complexe de quatre virus. Deux d’entre eux appartiennent au genre des polérovirus, ils sont génétiquement proches et responsables des symptômes de la jaunisse modérée : le Beet Mild Yellowing Virus (BMYV) et le Beet Chlorosis Virus (BChV). Le Beet Yellows Virus (BYV), responsable des symptômes de la jaunisse grave, est une espèce plus éloignée qui appartient au genre des clostérovirus. Le virus de la mosaïque (BtMV) appartient au genre des potyvirus. Ces virus sont exclusivement transmis par des pucerons vecteurs lorsqu’ils se nourrissent sur les feuilles de betterave, au premier rang duquel le puceron vert du pêcher Myzus persicae. La transmission est dite non-propagative car ces virus ne peuvent être transmis à la descendance des pucerons virulifères (porteurs du virus)(1) Au champ, les premiers symptômes apparaissent en moyenne entre 2 (30) et 6 semaines après l’inoculation. Le contrôle des pucerons doit donc s’effectuer bien avant l’apparition des symptômes.
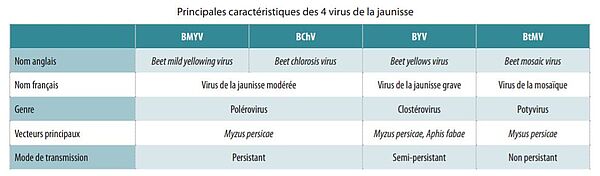
Le mode persistant repose sur une phase d’acquisition virale de 12 à 72 heures lors de l’alimentation sur une plante infectieuse. Ce délai est dû à la circulation des particules virales dans le corps du puceron : celles-ci doivent en effet passer dans la lumière du tube digestif, traverser la barrière épithéliale pour accéder à l’hémolymphe puis enfin migrer vers les cellules épithéliales des glandes salivaires du puceron. Les pucerons virulifères ne deviennent infectieux que lorsque les particules virales ont atteint la lumière des glandes salivaires. Les polérovirus de la betterave sont par ailleurs non-multipliants, c’est-à-dire qu’ils ne se reproduisent pas dans le corps du puceron, qui est un simple vecteur.
Il est à noter que les polérovirus n’infectent que les vaisseaux du phloème au sein des feuilles, que le puceron doit donc atteindre pour acquérir le virus. C’est aussi pour cette raison que la transmission mécanique de plante à plante est pratiquement impossible même en cas de blessure. La persistance correspond au fait que les pucerons demeurent infectieux jusqu’à épuisement du stock de particules virales, après plusieurs jours voire plusieurs semaines, soit généralement durant toute leur vie.(1)
Le mode semi-persistant repose sur une phase d’acquisition virale beaucoup plus rapide, de quelques minutes à quelques heures. En revanche, le virus ne se conserve qu’au plus 48h au niveau des pièces buccales de l’insecte, les taux de transmission étant déjà pratiquement nuls après 24h.(2) Il peut être retransmis dès l’acquisition mais il est en revanche perdu avec la mue du puceron. Les clostérovirus peuvent infecter tous les tissus de la feuille et pas seulement les vaisseaux du phloème, mais des expériences ont montré que la transmission mécanique de plante à plante était néanmoins très difficile à obtenir au laboratoire et vraisemblablement inexistante au champ.
Le mode non persistant repose sur une phase d’acquisition encore plus rapide. Quelques secondes suffisent pour que le puceron acquiert le virus et il peut être retransmis dès l’acquisition. Du fait de son temps d’acquisition très court, des pucerons qui effectuent des piqûres d’essais mais qui finalement ne s’alimentent pas peuvent acquérir le virus. Le virus se conserve quelques minutes à quelques heures au niveau des pièces buccales du puceron et n’est ni transmis à la descendance, ni conservé pendant la mue. La capacité du puceron à transmettre le virus diminue s’il touche des surfaces avec son stylet ou à mesure qu’il effectue des piqûres d’essais.(10)
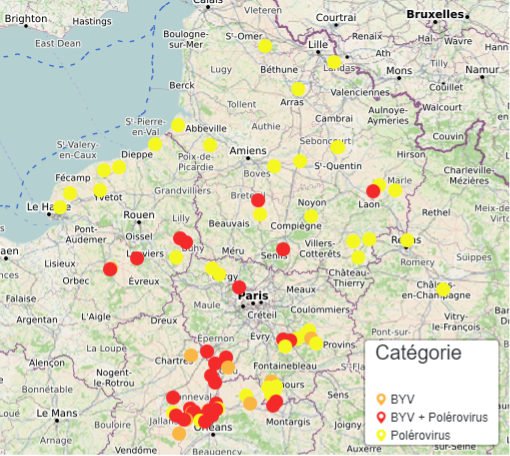
La prévalence des différents virus de jaunisse est très variable d’une année à l’autre : 2019 était caractérisée par une forte prédominance des polérovirus en mono infection, 2020 par la présence de co-infections sur tout le territoire betteravier et 2021 par une situation contrastée régionalement (la région Centre Val-de-Loire était majoritairement touchée par des co-infections BYV/polérovirus alors que sur le reste du territoire des polérovirus seuls ont majoritairement été détectés).
Prévalence virale en 2021
En région Centre-Val-de-Loire, la région la plus touchée par la jaunisse, 84 % des échantillons contenaient du BYV et 78 % des échantillons contiennent des polérovirus. 62% étaient co-infectés par le BYV et au moins un polérovirus.
Dans les autres régions 19% des échantillons étaient co-infectés par le BYV. Seulement 6% des échantillons étaient positifs au BYV seul et 75 % des échantillons étaient infectés par au moins un polérovirus.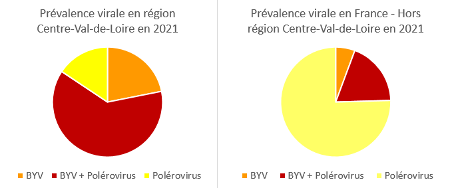
Prévalence virale en 2021 obtenue par tests sérologiques (Elisa)
Prévalence virale en 2020
En 2020, en situation de forte pression jaunisse, des analyses virales réalisées par l’ITB sur plus de 160 parcelles ont montré que tous les virus sont largement présents et que les multi-infections étaient très fréquentes : 72 % d’infections par 2 familles de virus ou plus et 45 % d’infections par les trois familles de virus.
82 % des betteraves symptomatiques étaient infectées par des polérovirus responsables de la jaunisse modérée, 70 % par le BYV et 64 % étaient infectées par le BtMV.
Prévalence virale en 2019
En 2019, les analyses virales réalisées dans le cadre du projet Casdar ExTraPol, en collaboration avec l'INRAE de Colmar et le GEVES, ont montré une très forte prévalence des polérovirus responsables de la jaunisse modérée (93 %) en France dont 72 % de BChV, contre seulement 7 % de BYV et une faible proportion de co-infections (4 %).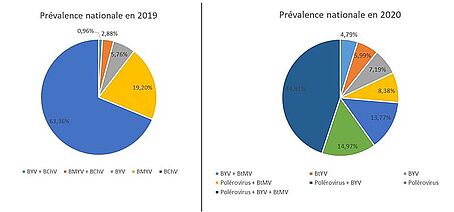
prévalence virale en 2019 et 2020 obtenue par test moléculaire (PCR)
Au moins 200 espèces de plantes appartenant à une vingtaine de familles botaniques sont hôtes de virus de jaunisses. Parmi ces espèces, certaines ne sont pas ou peu présentes en France (espèces originaires du continent européen, espèces ornementales, ...). D’autres plantes sont en revanche très communes, par exemple la capselle (Capsella bursa-pastoris) (27) (23) qui est hôte du BMYV ou le lamier pourpre (Lamium purpureum) (21) (24) qui est hôte du BYV et de polérovirus. Au total une cinquantaine de plantes (liste disponible dans le tableau ci-dessous) communes en France sont des hôtes connus d’au moins un virus de jaunisse de la betterave.
Les espèces cultivées et hôtes des virus de la jaunisse de la betterave appartiennent à la famille des Chenopodiaceae, notamment les épinards qui sont hôtes du BYV, BChV, BMYV et BtMV (21) (27)(23) (25) (26). Les betteraves hivernantes (betteraves porte graines, …) sont identifiées comme une source importante de réservoir viral (22)
Des adventices couramment présentes dans les parcelles de betterave sont également hôtes de virus de jaunisses comme le mouron blanc (Stellaria media) (21) (23) (24) (26) (28), qui est à la fois hôte des virus de la jaunisse modérée et du BYV, les amarantes et les chénopodes.
Nom commun | Nom Latin | Famille |
Ansérine | Argentina anserina (L.) Rydb. | Rosaceae |
Armoise commune | Artemisia vulgaris | Asteraceae |
Arroche étalée | Atriplex patula L. | Amaranthaceae |
Arroche couchée | Atriplex hastata L. | Amaranthaceae |
Betterave | Beta vulgaris | Amaranthaceae |
Betterave à gros fruits | Beta vulgaris ssp macrocarpa | Amaranthaceae |
Betterave maritime | Beta vulgaris ssp maritima | Amaranthaceae |
Epinard-fraise | Blitum capitatum L. | Amaranthaceae |
Souci des champs | Calendula arvensis L. | Asteraceae |
Caméline | Camelina sativa Crantz. | Brassicaceae |
Bourse-à-pasteur | Capsella bursa-pastoris (L.) Medik. | Brassicaceae |
Ansérine hybride ; Chénopode à feuilles de stramoine | Chenopodiastrum hybridum (L.) S.Fuentes, Uotila & Borsch | Amaranthaceae |
Chénopode blanc | Chenopodium album L. | Amaranthaceae |
Chénopodes | Chenopodium sp. | Amaranthaceae |
Chénopode fétide | Chenopodium vulvaria L. | Amaranthaceae |
Chrysanthème des moissons | Chrysanthemum segetum | Asteraceae |
Cirse des champs | Cirsium arvense (L.) Scop. | Asteraceae |
Claytone de Cuba | Claytonia pertoliata Donn ex Willd. | Montiaceae |
Crépide verdâtre | Crepis capillaris | Asteraceae |
Bec-de-grue à feuilles de ciguë | Erodium cicutarium (L.) L'Hér. | Geraniaceae |
Euphorbe réveille-matin | Euphorbia helioscopia L. | Euphorbiaceae |
Renouée faux liseron | Fallopia convolvulus (L.) Á.Löve | Polygonaceae |
Fumeterre officinale | Fumaria officinalis L. | Papaveraceae |
Chrysanthème des blés ; Chrysanthème des moissons | Glebionis segetum (L.) Fourr. | Asteraceae |
Lamier à feuilles embrassantes | Lamium amplexicaule L. | Lamiaceae |
Lamier pourpre | Lamium purpureum L. | Lamiaceae |
Grageline | Lapsana communis L. | Asteraceae |
Bourse-de-Judas | Lepidium campestre (L.) R.Br. | Brassicaceae |
Passerage drave | Lepidium draba L. | Brassicaceae |
Chénopode à graines nombreuses | Lipandra polysperma (L.) S.Fuentes, Uotila & Borsch | Amaranthaceae |
Mouron rouge | Lysimachia arvensis (L.) U.Manns & Anderb. | Primulaceae |
Camomille vraie | Matricaria chamomilla | Asteraceae |
Mercuriale annuelle | Mercurialis annua L. | Euphorbiaceae |
Coquelicot douteux ; Petit Coquelicot | Papaver dubium L. | Papaveraceae |
Coquelicot | Papaver rhoeas L. | Papaveraceae |
Phacélie à feuilles de tanaisie | Phacelia tanacetifolia Benth. | Boraginaceae |
Plantain lancéolé | Plantago lanceolata L. | Plantaginaceae |
Grand plantain | Plantago major L. | Plantaginaceae |
Pourpier maraîcher | Portulaca oleracea L. | Portulacaceae |
Séneçon commun | Senecio vulgaris L. | Asteraceae |
Moutarde blanche | Sinapis alba L. | Brassicaceae |
Jotte ; Moutarde des champs | Sinapis arvensis L. | Brassicaceae |
Laiterons | Sonchus sp. | Asteraceae |
Spargoute des champs | Spergula arvensis L. | Caryophyllaceae |
Mouron blanc ; Stellaire intermédiaire | Stellaria media (L.) Vill. | Caryophyllaceae |
- | Taraxacum sp. | Asteraceae |
Monnoyère | Thlaspi arvense L | Brassicaceae |
Véronique petit-chêne | Veronica chamaedrys L. | Plantaginaceae |
Véronique commune | Veronica persica Poir. | Plantaginaceae |
- | Veronica sp. | Plantaginaceae |
Pensée des champs | Viola arvensis Murray | Violaceae |
Pensée sauvage ; Pensée tricolore | Viola tricolor L. | Violaceae |
Liste des espèces communes en France et hôte d’au moins un des virus responsables de la jaunisse de la betterave (21)(23)(24)(25)(26)(27)(28)
Des travaux complémentaires seront menés dans le cadre du PNRI par INRAE afin de savoir, parmi les plantes hôtes, celles sur lesquelles les pucerons vecteurs de jaunisse se nourrissent avant de migrer sur betterave. Ils permettront de mieux comprendre quels sont les réservoirs de virus effectifs sur le terrain.
Les pucerons sont des insectes de la famille des Aphididae. En Europe, il en existe environ 900 espèces dont quelques dizaines sont des ravageurs des cultures. Ils passent l’essentiel de l’année sous un mode de reproduction asexué. Les larves deviennent adultes en moins de 15 jours entre 15 et 20°C ce qui fait qu’une quinzaine de générations se succèdent entre le printemps et l’automne. Ce type de reproduction s’effectue sur les hôtes secondaires des pucerons, principalement des plantes herbacées dont les fabacées et les chénopodiacées. A l’automne, la baisse de la photopériode induit un second mode de reproduction sexué sur leurs hôtes primaires qui permet un brassage génétique au sein de l’espèce. Ce cycle complet avec alternance d’une phase sexuée et asexuée est dit holocyclie. Cependant, la douceur hivernale a favorisé l’absence de phase sexuée, on parle alors d’anholocyclie. Les espèces de pucerons qui alternent leur cycle entre hôtes primaires et secondaires sont dites diœciques. Myzus persicae et Aphis fabae sont des espèces holocycliques diœciques mais qui peuvent être anholocycliques dans les régions à climat doux(3). Il existe 4 stades larvaires successifs qui s’achèvent en 8 à 10 jours avant la mue imaginale qui produira l’adulte. Un adulte est capable de vivre jusqu’à 120 jours et donne naissance en moyenne à 40 à 60 larves au cours de sa vie.
Morphologie
Les adultes mesurent entre 1,2 et 2,5 mm. Les aptères sont vert clair à vert jaunâtre avec, au sein de la colonie, des individus rosés qui sont les futurs ailés (nymphes). Les ailés sont vert clair avec une plaque sombre sur l'abdomen échancrée latéralement et perforée.(3)
Cycle biologique
Lorsque la reproduction est holocyclique, les œufs d’hiver sont pondus en automne à l’aisselle des écailles de bourgeons de pêchers ou de cerisiers à grappes. Ces œufs sont très résistants au froid et leur limite de répartition dans les régions froides est l’aire de répartition des hôtes primaires. Ils colonisent la face inférieure des feuilles en produisant de manière clonale plusieurs générations successives d’individus aptères au printemps puis des individus ailés rejoignent les hôtes secondaires de la dissémination entre parcelles. Les individus sexués ailés n’apparaissent qu’à l’automne lors des vols de remigration vers les hôtes primaires.(4)
Dans les climats tempérés comme en France, Myzus persicae peut rester sous forme asexuée toute l’année. Les adultes et les larves peuvent survivre en hiver sur leurs hôtes secondaires : crucifères, épinards d’hivers, diverses adventices, ainsi que dans les silos de betteraves fourragères et sucrières, sur les betteraves potagères conservées avec leurs repousses, ou sur des repousses de betteraves abandonnées à la récolte. La météo hivernale à un effet sur le développement et la survie des adultes et des larves hivernants de Myzus persicae : on estime que la température à partir duquel les nymphes issues de la reproduction asexuée se développent est de 4°C.(11) En Angleterre, la date des premiers vols est corrélée positivement aux températures moyennes en janvier et février. Le taux de mortalité des larves et des adultes dépend également de la météo en hiver. La résistance au froid de M. persicae dépend de la souche mais aussi de l’acclimatation des pucerons.(12)(13)(14)
Les adultes produits par parthénogénèse peuvent être ailés ou aptères. Le pourcentage de pucerons ailés augmente lorsque la densité de pucerons augmente et lorsque la qualité nutritionnelle de la plante hôte diminue.
Le taux de reproduction, le temps nécessaire à la larve pour devenir adulte et l’espérance de vie du puceron dépendent des conditions météorologiques mais aussi de la plante hébergeant le puceron. A une température constante de 23 °C, les clones mettent 6,5 à 7 jours pour devenir adulte. L’espérance de vie une fois adulte varie de 15 à 25 jours selon la plante hôte. Sur des hôtes favorables à 23°C, une femelle produit 70 à 80 nymphes durant sa vie.(15) Lorsque la température diminue la durée de développement diminue : il faut ainsi environ 14 jours à 15°C pour que les pucerons deviennent adulte et plus de 60 jours à 5°C. Le nombre de larves produites par jour diminue également avec la température et tombe à 1,1 œuf par jours à 10 °C.(16)
Nuisibilité
Nuisibilité directe
Myzus persicae n’occasionne que très rarement des dégâts directs sur betterave.
Capacités de transmission virale
Selon différentes études conduites en laboratoire, Myzus persicae est le vecteur le plus efficace des trois virus de la jaunisse avec des taux de transmission compris entre 28 et 100 % pour les polérovirus (BMYV et BChV) et entre 51 et 73 % pour le BYV.(2)(5)(6)(7) Il est également capable de transmettre le BtMV mais les taux de transmission au champ ne sont pas connus. De plus, au niveau épidémiologique, les pucerons verts ailés seraient les principaux responsables de la dissémination virale au champ car ils sont beaucoup plus mobiles que les ailés noirs, sédentaires, et dont la contribution serait en réalité négligeable.(8)

Morphologie
Les adultes mesurent entre 1,5 et 2,6 mm. Les aptères sont trapus, de couleur noir mat à verdâtre avec au sein des colonies, des individus ayant des bandes cireuses blanches sur l’abdomen (nymphes). Les ailés sont de couleur sombre avec de courtes antennes.(3)
Cycle biologique
Les œufs d’hiver sont pondus à l’aisselle des écailles de bourgeons du fusain d’Europe ou de la viorne obier. Les femelles fondatrices issues des œufs d’hiver produisent une génération de femelles aptères qui produisent une génération de femelles ailées, le tout de manière asexuée. Les femelles fondatrices colonisent les parcelles contenant leurs hôtes secondaires et produisent des colonies d’aptères sur la face inférieure des feuilles. Une nouvelle génération d’individus sexués apparaît à l’automne et retourne sur les hôtes primaires afin d’y pondre les œufs qui passeront l’hiver.(4)
Nuisibilité
Nuisibilité directe
Les colonies de puceron noir peuvent occasionner des dégâts directs en s’alimentant sur les feuilles de betterave (perte de vigueur des plantes). Ces pucerons produisent également du miellat à l’origine du développement de la fumagine, c’est-à-dire de moisissures noires sur la face inférieure des feuilles. L’impact sur le rendement in fine est en cours d’évaluation à l’ITB.
Capacités de transmission virale
Selon les mêmes études précédemment citées, les capacités de transmission d’Aphis fabae vis-à-vis des virus de la jaunisse modérée (BMYV et BChV) sont pratiquement nulles, quels que soient les isolats viraux et clones de pucerons testés.(5)(6)(7) En revanche, Aphis fabae est un bon vecteur du virus de la jaunisse grave, avec des taux de transmission compris entre 28 et 40 %, et un vecteur du BtMV. Mais Myzus persicae reste tout de même un meilleur vecteur que Aphis fabae.(2)


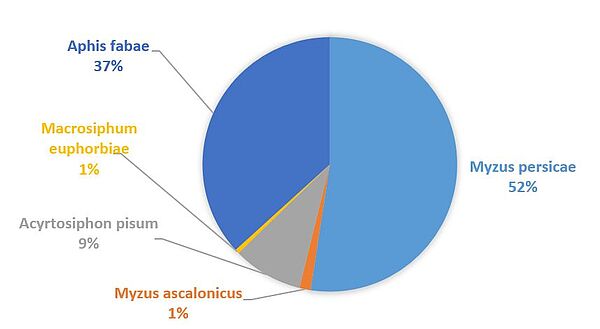
Il existe d’autres espèces de pucerons vecteurs des virus de la jaunisse telles que Macrosiphum euphorbiae, Acyrtosiphon pisum, Myzus ascalonicus ou Aulacorthum solani pour n’en citer que quelques-unes mais celles-ci sont minoritaires et leurs capacités de transmission bien plus faibles que celles de Myzus persicae (6) pour le BYV et les polérovirus responsables de la jaunisse modérée. En revanche la transmission de potyvirus par des espèces de pucerons non hôtes a été reporté dans la littérature sur plusieurs complexes hôtes/virus/pucerons, et Acyrthsiphon pisum, Macrosiphum euphorbiae ou Rhopalosiphum padi ont la capacité de transmettre le BtMV.(18)(19) A titre d’exemple, voici la répartition globale des captures d’individus ailés de 5 espèces différentes dans un réseau de cuvettes jaunes relevées de manière hebdomadaire au printemps 2019.
La jaunisse se manifeste au champ par l’apparition à partir de fin juin de foyers (ou ronds) jaunes bien distincts au sein d’une parcelle. En fin d’été ou à l’automne, si la parcelle est très infectée, les foyers peuvent se rejoindre et la maladie prendre un aspect plus diffus.

Les symptômes débutent par un jaunissement diffus envahissant progressivement la feuille à partir de son sommet, mais laissant les nervures vertes. La feuille s’épaissit, devient cassante et prend peut prendre une teinte orangée. A un stade plus avancé, ces taches virent au rouge-brun et peuvent se rejoindre de sorte que la teinte rougeâtre peut devenir dominante.(5)
Plus tardivement, lorsque la maladie est bien développée, des attaques du champignon Alternaria tenuis peuvent se produire, provoquant une nécrose s’étendant à partir des bords de la feuille.(5)
Il n’est pas aisé de faire la différence à l’œil nu entre les symptômes de jaunisse modérée et ceux de jaunisse grave, d’autant plus qu’une même plante peut potentiellement être co-infectée par plusieurs virus. Des analyses en laboratoires après prélèvement de feuilles permettent de déterminer les virus impliqués.
Les premiers symptômes sont l’apparition de minuscules points clairs sur le limbe, puis d’un éclaircissement des nervures secondaires (ou gravure des feuilles, qui a donné son nom à la maladie) et enfin le développement de taches jaune-citron.4 A un stade plus avancé, ces taches virent au rouge-brun et peuvent se rejoindre de sorte que la teinte rougeâtre peut devenir dominante.5

Chez une betterave infectée, les nervures des feuilles du cœur s'éclaircissent et tendent à devenir blanches. Sur les feuilles plus âgées, on peut observer des marbrures d'un vert plus clair.
Il n’est pas aisé de faire la différence à l’œil nu entre les symptômes de jaunisse modérée et ceux de jaunisse grave, d’autant plus qu’une même plante peut potentiellement être co-infectée par plusieurs virus. Des analyses en laboratoires après prélèvement de feuilles permettent de déterminer les virus impliqués.

Symptômes foliaire de mosaïque de la betterave.
Ne pas confondre !
Le jaunissement des feuilles peut être également dû à certain nombre de facteurs dont une déficience physiologique (excès ou une carence en manganèse, carence en magnésium, carence en bore), un stress hydrique, une mauvaise structure du sol, l’attaque d’autres bioagresseurs, etc. Cependant, la spécificité des symptômes de jaunisse réside dans l’épaississement et la teinte jaune orangée prise par les feuilles atteintes, ainsi que dans l’apparition de foyers primaires irrégulièrement répartis au sein d’une parcelle.
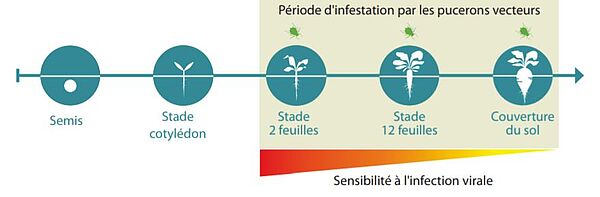
Stades de sensibilité
La période à risque commence dès l’apparition des premiers pucerons dans les parcelles au plus tôt fin avril début mai, soit à partir du stade 2 feuilles jusqu’à la couverture du sol fin juin. Sur des plantes non protégées, la dissémination du virus et des symptômes associés peut continuer jusqu’à la récolte, mais les plantes adultes (au-delà du stade 12 feuilles) sont naturellement plus résistantes aux pucerons et à la transmission virale. Ce phénomène est connu sous le nom de "résistance à maturité", il impacte notamment la capacité des pucerons à se nourrir et se multiplier sur les plantes, mais ses déterminants sont à ce jour encore mal connus.
Chaque année, de 200 à 300 parcelles sont suivies chaque semaine dans le cadre du BSV (Bulletin de santé du végétal), sur lesquelles des comptages d’aptères dans les betteraves sont effectués au printemps. L’ITB a défini à partir de son expertise historique un seuil de risque égal à 10 % de betteraves colonisées par au moins un aptère vert.
Cette analyse de risque est complétée par des conseils de gestion dans les notes d'informations régionales qui sont communiquées aux planteurs de manière hebdomadaire sur le site internet de l'ITB et par mail pour les abonnés.
Depuis 2019, « Alerte pucerons » est disponible en ligne sur le site de l’ITB. Il s’agit d’une carte interactive qui présente 4 niveaux de seuils (non atteint, seuil de risque du premier traitement atteint, seuil de risque du second traitement atteint, seuil de risque du troisième traitement atteint) établis à partir des observations du BSV et mis à jour quotidiennement tout au long de la période de sensibilité à la jaunisse (de mi-avril à fin juin).
Les données que nous avons acquises depuis 2017 montrent que sur des parcelles infectées par la jaunisse modérée, les pertes de productivité dans les ronds de jaunisse se situent en moyenne autour de 25 %. A l’échelle de la parcelle, nous estimons que 10 % de surface atteinte entraîne une perte de rendement d’environ 3 t/ha.
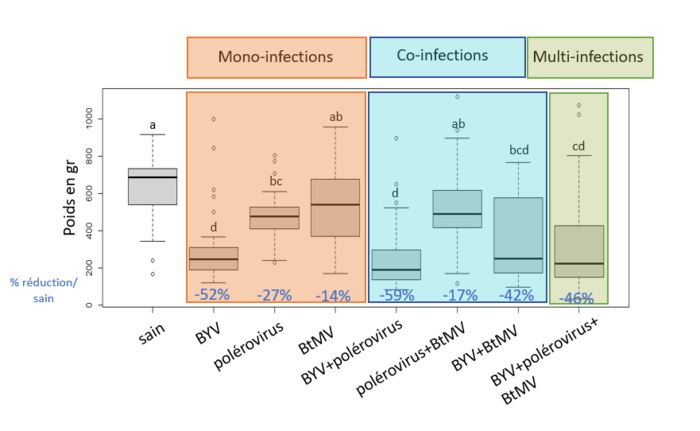
Une étude menée dans le cadre d’une collaboration entre l’ITB et INRAE Colmar dans le cadre du PNRI en 2021, en situation d’inoculation artificielle a mis en évidence des pertes de rendements que les pertes de rendement dépendent du virus inoculé : le BYV a occasionné une diminution du poids des betteraves de 52% et une diminution de la richesse de 4% (soit - 0,7 point de richesse). Les polérovirus ont entrainé une diminution du poids de racine de 27%. Les betteraves inoculées mécaniquement avec le BtMV tendent à avoir un poids de racine plus faible que les betteraves saines ( -14% de poids racine) sans que cette différence soit significative. On observe en revanche une augmentation de la richesse de 6% pour ces betteraves inoculées avec le BtMV.
Cette étude n’a pas permis de montrer d’augmentation des pertes de rendements en situation de co-infection. Les pertes de rendement sont statistiquement similaires aux pertes causées par le virus le plus dommageable seul.
Selon la littérature grise le BtMV causerait également une diminution importante de la faculté des graines à germer.
Des études menées au Royaume-Uni montrent que les pertes de rendement sont d’autant plus importantes que l’infection survient à un stade précoce du développement de la betterave. (21) (29)
Le Teppeki (50 % de flonicamide) est homologué depuis le 21 décembre 2018 avec les conditions d’emploi suivantes :
Le Karaté K (lambda-cyhalothrine 5 g/L et pirimicarbe 100 g/L) est également homologué sur betteraves contre les pucerons depuis 2011, mais son utilisation est fortement déconseillée car les populations de Myzus persicae y sont largement résistantes et car ce produit n’est pas sélectif vis-à-vis des auxiliaires. Des applications avec ce produit peuvent donc accentuer le degré d’infection par la jaunisse sur une parcelle, malgré son efficacité contre les pucerons noirs.
Des projets collaboratifs de recherches sont en cours pour mieux connaitre les virus responsables de la jaunisse ainsi que les pucerons vecteurs, développer de nouvelles méthodes pour prévenir les pertes de rendements causées par les jaunisses, et lutter contre les pucerons vecteurs de jaunisse. L’ITB prend part à 3 de ces projets :
- Le projet Casdar ExTraPol (2019-2022)
- Le projet Modefy (2021-2024)
- Le Plan National de Recherche et d’Innovation (PNRI) vers des solutions opérationnelles contre la jaunisse de la betterave.
Références
|
Références
|

![]()
Agrément conseil de l’ITB à l’utilisation des produits phytosanitaires n° 7500002.
Assurance RC n° 05421646Y/1025.
Le portail EcophytoPIC recense les techniques alternatives à l’utilisation des produits phytopharmaceutiques.
Organisme agréé Crédit d'impôt Recherche
L'Institut Technique de la Betterave est
membre du réseau Acta
Institut Technique Agricole Qualifié
par le Ministère de l’Agriculture et de l’Alimentation
