
La cercosporiose est une maladie fongique foliaire estivale qui peut apparaître dès le mois de juin. Elle est une des maladies foliaires les plus destructrices menaçant la production mondiale de betterave sucrière. Les surfaces concernées par la cercosporiose sont en augmentation sur tout le territoire. Environ 300 000 ha de betteraves sont aujourd’hui concernés par le développement de la cercosporiose à l’échelle nationale. Aujourd’hui, aucune région n’est épargnée. Les zones les plus touchées sont l’Alsace, la Champagne, l’Ile-de-France et la région Centre.
Cercospora beticola, l’agent responsable de la cercosporiose, a été décrit comme une espèce du genre Cercospora en 1876. C. beticola est probablement originaire d’Europe centrale et du bassin méditerranéen (Groenewald et al., 2005). Les premiers cas de dommages liés à la cercosporiose ont été rapportés dès 1878 en Europe. En France, la maladie a été décrite pour la première fois en 1887, elle est décrite comme un problème aux Etats-Unis à partir de 1895. Aujourd’hui, c’est plus d’un tiers de la surface mondiale de production de betterave sucrière qui est affectée (Tan et al., 2023).
Dernière mise à jour : janvier 2026
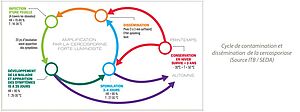
La cercosporiose est causée par le champignon Cercospora beticola, un ascomycète de la famille des Mycosphaerellaceae. Ce champignon a un modèle de développement hémibiotrophe : son cycle se compose d’une phase biotrophique asymptomatique pendant l’incubation puis transitionne vers un mode nécrotrophe entrainant la mort des cellules infectées pour se nourrir (Oerke et Steiner, 2025). La cercosporiose est caractérisée par des cycles répétés de contamination (germination des conidies et infection de l’hôte) et dissémination (sporulation des conidies, détachement et dissémination à la surface des plantes) (Franc, 2010) :
Le développement de la cercosporiose au champ suit donc un processus polycyclique caractérisé par une chaîne de cycles d’infections ininterrompue pendant l’été si les conditions sont favorables (Rossi, 2000).
C. beticola peut contaminer une grande variété de plantes appartenant aux familles des chénopodiacées (betterave potagère, bette, épinard), des acanthacées, des apiacées (céleri), des astéracées, des brassicacées, des malvacées, des plumbaginacées ou encore des polygonacées (Groenewald et al., 2006).
C. beticola se conserve principalement sur les résidus de récolte infestés : feuilles, collets ou tas de déterrage. Les conidies peuvent survivre jusqu’à 8 mois sur ces résidus. Cependant, l'inoculum peut se conserver bien plus longtemps, jusqu’à 3 ans, au sein d’une structure de conservation appelée pseudostromate. Cette structure contient des hyphes et des conidiophores qui peuvent sporuler en conditions favorables. Les résidus de récolte constituent donc le réservoir d’inoculum principal de C. beticola qu’il convient alors de détruire et d’enfouir afin d’interrompre le cycle de la maladie. Les graines contaminées et les autres plantes hôtes peuvent constituer des sources secondaires d’infection (Franc, 2010). Il a de plus été montré dans des essais au champ aux États-Unis que la profondeur d’enfouissement de l’inoculum de C. beticola affecte sa durée de conservation (Khan et al., 2008). En effet, 10 mois après inoculation à 0, 10 et 20 cm de profondeur, la viabilité de l’inoculum était réduite de 59, 74 et 76 % respectivement. Après 22 mois, l’inoculum n’a pas survécu lorsqu’il était enterré à 10 ou 20 cm de profondeur, alors qu’il était encore partiellement viable à la surface du sol. L’enfouissement profond des résidus de récolte doit donc permettre de réduire la viabilité de l’inoculum de Cercospora beticola dans le sol et ainsi ralentir le développement de la maladie.
Lors de sa phase nécrotrophe, C. beticola produit des toxines photosensibles telles que la cercosporine, activée par la lumière visible et proche UV. Elle n’est pas spécifique à une plante hôte et est produite par la plupart des espèces de champignons du genre Cercospora (Daub et Ehrenshaft, 2000). Dans le cas de C. beticola, elle constitue un facteur de virulence mais n’est pas nécessaire au succès du pathogène. La cercosporine activée par la lumière, en réaction directe ou indirecte avec l’oxygène, produit des composés ROS, espèces réactives de l’oxygène. À haute concentration, ces composés causent des dommages aux cellules de l’hôte (Birben et al., 2012). Le champignon est protégé de ses propres toxines par des mécanismes d’auto-résistance : exportation des toxines via des transporteurs ou détoxification. La béticoline est une autre toxine responsable de la virulence de C. beticola.
Diversité génétique et reproduction
C. beticola a longtemps été considérée comme appartenant au complexe Cercospora apii dont l’hôte est le céleri. Des analyses moléculaires ont cependant révélé qu’il s’agit bien d’espèces distinctes l’une de l’autre (Groenewald et al., 2005). La reproduction sexuée chez C. beticola n’a pas clairement été démontrée mais la grande diversité génétique du champignon est une preuve indirecte. La variabilité génétique des populations de C. beticola a été étudiée dans un projet coordonné par l’ITB de 2021 à 2024, en collaboration avec l’INRAE, l’ANSES et grâce à la contribution financière du MAASA. Les résultats montrent que la variabilité génétique en France est forte, il n’existe pas de souche dominante, ni de structuration géographique des souches présentes sur le territoire.
La maladie apparaît généralement à partir du mois de juillet, parfois dès le mois de juin, sur la face supérieure des feuilles primaires donc extérieures du bouquet foliaire. Elle se développe sous la forme de taches circulaires grisâtres et déprimées, de 3 à 5 mm de diamètre à maturité, souvent entourées d'une bordure rougeâtre (Windels et al., 1998). Si les conditions sont favorables au développement du champignon, les taches se densifient et fusionnent, les phytotoxines s’accumulent, entraînant le jaunissement puis le dessèchement complet des feuilles touchées. Les feuilles intérieures sont ensuite touchées à leur tour. La mortalité des feuilles peut donner un aspect de "champ brûlé" aux parcelles fortement touchées. La destruction du bouquet foliaire induit une forte repousse de feuilles destinée à maintenir la capacité photosynthétique de la plante si les conditions climatiques ne sont pas trop sèches.

Ne pas confondre !
Il est possible de confondre la cercosporiose avec la ramulariose ou encore la bactériose à Pseudomonas. Il faut ainsi s’assurer à la loupe de la présence de conidiophores (granulation noire) au centre de la tâche.
La cercosporiose entraîne une diminution importante du saccharose, pouvant atteindre jusqu’à 50 % en cas de forte pression maladie non maîtrisée (Shane et Teng, 1992). La plante compense la destruction foliaire par une repousse de feuilles au détriment des réserves en sucres de la racine.
La maladie provoque également une augmentation des impuretés dans la racine compliquant le processus de récupération du saccharose et entrainant des coûts de transformation plus élevés (Shane et Teng, 1992).
Enfin, lors du stockage en silo, les plantes malades sont plus sensibles à la pourriture racinaire, ce qui augmente les pertes post-récolte (Smith, 1971).
L’apparition et le développement de la cercosporiose dépendent de différents facteurs :
Les principaux facteurs de risque sont listés dans le tableau ci-dessous :

Les moyens de lutte reposent sur une combinaison de leviers répondant aux objectifs de la protection intégrée : allongement des rotations, enfouissement profond des résidus de récolte, implantation de variétés tolérantes ou résistantes, application de produits fongicides en végétation.
Compte-tenu de l’importance de l’inoculum primaire dans la dissémination de la maladie, l’allongement des rotations avec des plantes non-hôtes et l’enfouissement profond des résidus de récolte sont indispensables pour réduire la quantité d’inoculum de C. beticola dans le sol. Il est également judicieux de ne pas emblaver en betteraves à proximité d’une parcelle sévèrement contaminée l’année précédente.
Adapter le choix variétal dans les zones à risque
À l’échelle de la parcelle, il est important d’adapter le choix variétal au niveau de risque encouru (voir facteurs de risques plus haut) ainsi qu’à la date de récolte. Le semis d’une variété sensible ne doit avoir lieu qu’en situation de risque faible (pas de proximité avec des foyers infectieux, zone non irriguée) et/ou pour des récoltes précoces.
Les variétés résistantes à la cercosporiose montrent une progression plus lente de la sévérité comparées aux variétés sensibles. Ainsi, le vol de spores est significativement plus faible dans les parcelles de variétés résistantes, ce qui réduit le niveau d’inoculum présent (Yang et al., 2025).
À l’heure actuelle, les variétés résistantes à la cercosporiose ne possèdent qu’une résistance partielle qui ne permet pas de contrôler à elle seule le développement de la cercosporiose, des applications phytosanitaires complémentaires restent nécessaires (Rangel et al., 2020 ; Yang et al., 2025).
La résistance variétale est monogénique (due à un gène majeur de résistance) et/ou polygénique (due à plusieurs gènes mineurs) selon la variété. Dans le cas d’une résistance monogénique, les gènes majeurs (par exemple BvCR4) fonctionnent selon le modèle “gène de résistance – gène d’avirulence”. Le pathogène C. beticola sécrète un effecteur d’avirulence (par exemple AvrCR4). La protéine codée par le gène de résistance dans la plante reconnaît cet effecteur et déclenche des réponses de défense empêchant la propagation de l’infection. En revanche, la résistance peut être contournée par des populations de champignons évolutives présentant une délétion ou mutation de l’effecteur, devenant virulentes sur les variétés jusque-là résistantes. Ce phénomène de contournement de résistance a été observé aux États-Unis pour le gène de résistance BvCR4 et le gène d’avirulence associé AvrCR4 (Chen et al., 2024). Ces observations récentes suggèrent que C. beticola évolue rapidement. Cette évolution peut être accélérée par la sélection imposée par le déploiement de variétés résistantes monogéniques, ce qui contraint les sélectionneurs à privilégier les variétés à résistance polygénique, plus difficilement contournable.
Le développement de la cercosporiose peut être limité par l’application de fongicides de contact ou systémiques. Les produits de contact ont une action préventive, ils agissent à la surface des plantes en empêchant la germination des spores avant qu'ils ne pénètrent dans les tissus. Les produits systémiques pénètrent dans la plante et se déplacent à travers les tissus par la sève, c’est une action préventive parfois complétée par une action curative contre les infections déjà établies. Lorsque certains éléments minéraux (tels que le cuivre) sont pulvérisés en mélange avec des fongicides, on observe une augmentation significative de l’activité de la catalase, de la peroxydase et de la polyphénoloxydase chez la betterave sucrière, ce qui atténue les dommages cellulaires.
L’efficacité des fongicides est diminuée par l'émergence de souches résistantes de C. beticola. Une résistance apparait après l'utilisation généralisée et répétée d'une même classe de fongicides (Rosenzweig et al., 2020). Le développement de résistance est favorisé par la nature polycyclique de ce champignon et son haut taux de sporulation, qui permet une sélection des souches lors de la saison, selon les produits fongicides utilisés. L'utilisation alternée de différentes classes de fongicides (donc de modes d’actions) permet de limiter la sélection de souches résistantes de C. beticola. Il est aussi recommandé d’appliquer les fongicides en mélange pour une plus grande efficacité et une réduction de la pression de sélection pour des isolats résistants (Hernandez et al., 2024). Ces méthodes permettent de prolonger l'efficacité des fongicides.
La résistance aux substances actives de la famille des strobilurines est aujourd’hui généralisée et, dans une moindre mesure, les triazoles sont aussi concernées. Seule l’utilisation de molécules multisites comme le cuivre évite l’apparition de souches résistantes, même si l’utilisation de cuivre n’est pas une solution durable à cause de son impact négatif sur la fertilité des sols.
Le suivi de ces résistances mené en collaboration entre l’ITB, l’INRAE et l’ANSES est important pour mettre en place des mesures de gestion de ces résistances.
Des produits de biocontrôle tels que des champignons ou des bactéries sont évalués par la recherche comme potentielles méthodes de lutte contre la cercosporiose. Au laboratoire, parmi 25 bactéries endophytes, Bacillus subtilis s’est montrée la plus efficace pour inhiber la croissance du pathogène (AbdElfatah et al., 2025). La bactérie a un effet antagoniste direct contre C. beticola (Sehsah et al., 2022). Les résultats en laboratoire n’ont pas encore été confirmés au champ. Plusieurs espèces de champignons de la famille des Trichoderma ont également montré une efficacité contre la cercosporiose, notamment par le déclenchement des réactions de défense de la plante (Kappel et al., 2022). D’autres produits tels que la chitosane, le bicarbonate, l’extrait de Moringa ou encore des huiles essentielles sont testés pour leurs effets de limitation de la croissance du pathogène ou d’induction de défenses de la plante (Kappel et al., 2022 ; Sehsah et al., 2022 ; Sehsah et al., 2023).
Surveiller l’évolution des maladies du feuillage dans les parcelles est important pour traiter au bon moment.
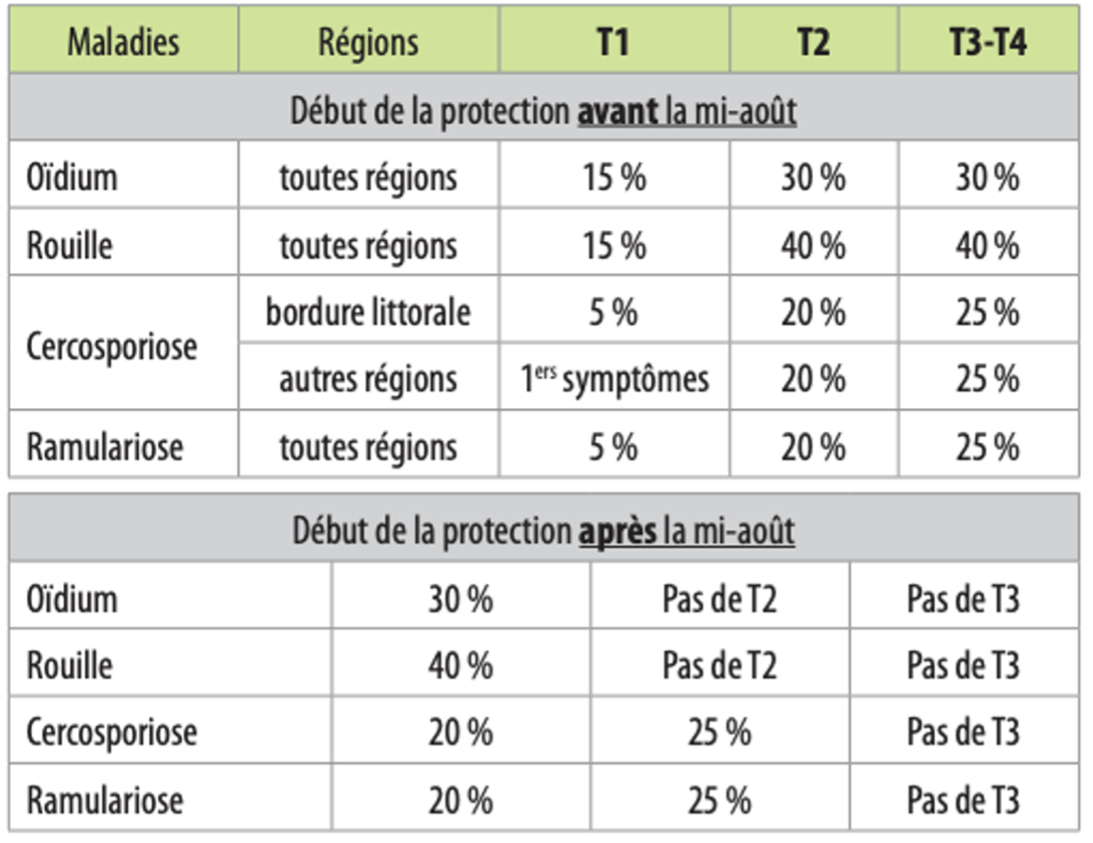
L’ITB a mis en place, depuis 2005, des seuils de déclenchement des traitements permettant de raisonner toute intervention fongicide.
Seuils de déclenchement des traitements aux maladies du feuillage (% de feuilles avec symptômes) :
L’OAD Alerte maladies est disponible
L'ITB met à disposition gratuitement la carte interactive "Alerte Maladies" afin de mieux accompagner les planteurs dans la lutte contre les maladies foliaires fongiques. Cet outil est mis à jour automatiquement à partir des notations hebdomadaires de l'ITB, des services techniques des sucreries et des autres observateurs du réseau de Suivi Biologique du Territoire. L’OAD est disponible à l’adresse suivante : http://maladies.itbfr.org/
En parallèle, l’ITB participe au développement de modèles épidémiologiques afin de mieux positionner les traitements. Des modèles de machine learning sont mis au point pour accompagner les producteurs dans la gestion de la maladie.
Références AbdElfatah, Heba-Alla, Hadeel M. M. Khalil Bagy, Mohamed S. Mohamed, Mennatallah M. A. El-Geddawy, Rofida F. Moftah, et Nashwa M. A. Sallam. « Biological Control of Cercospora Leaf Spot Disease of Sugar Beet Using Bacillus Subtilis ». Egyptian Journal of Biological Pest Control 35, no 1 (2025): 30. https://doi.org/10.1186/s41938-025-00866-z. Birben, Esra, Umit Murat Sahiner, Cansin Sackesen, Serpil Erzurum, et Omer Kalayci. « Oxidative Stress and Antioxidant Defense ». World Allergy Organization Journal 5, no 1 (2012): 9‑19. https://doi.org/10.1097/WOX.0b013e3182439613. Chen, Chen, Harald Keunecke, Felix Bemm, et al. « GWAS Reveals a Rapidly Evolving Candidate Avirulence Effector in the Cercospora Leaf Spot Pathogen ». Molecular Plant Pathology 25, no 1 (2024): e13407. https://doi.org/10.1111/mpp.13407. Daub, Margaret E., et Marilyn Ehrenshaft. « The Photoactivated CercosporaToxin Cercosporin : Contributions to Plant Disease and Fundamental Biology ». Annual Review of Phytopathology 38, no 1 (2000): 461‑90. https://doi.org/10.1146/annurev.phyto.38.1.461. Franc, Gary D. Ecology and Epidemiology of Cercospora Beticola. 2010. Groenewald, Marizeth, Johannes Z. Groenewald, et Pedro W. Crous. « Distinct Species Exist Within the Cercospora Apii Morphotype ». Phytopathology® 95, no 8 (2005): 951‑59. https://doi.org/10.1094/PHYTO-95-0951. Groenewald, Marizeth, Johannes Z. Groenewald, Thomas C. Harrington, Edwin C.A. Abeln, et Pedro W. Crous. « Mating Type Gene Analysis in Apparently Asexual Cercospora Species Is Suggestive of Cryptic Sex ». Fungal Genetics and Biology 43, no 12 (2006): 813‑25. https://doi.org/10.1016/j.fgb.2006.05.008. Hernandez, Alexandra P., Sarah Ruth, Christopher Bloomingdale, Linda E. Hanson, et Jaime F. Willbur. « Effects of Pyraclostrobin Rotations and Tank-Mixtures on Cercospora Beticola Sensitivity and Cercospora Leaf Spot Control in Sugarbeet ». Crop Protection 182 (août 2024): 106710. https://doi.org/10.1016/j.cropro.2024.106710. Kappel, Lisa, Nicole Kosa, et Sabine Gruber. « The Multilateral Efficacy of Chitosan and Trichoderma on Sugar Beet ». Journal of Fungi 8, no 2 (2022): 137. https://doi.org/10.3390/jof8020137. Khan, J., L. E. del Río, R. Nelson, et M. F. R. Khan. « Improving the Cercospora Leaf Spot Management Model for Sugar Beet in Minnesota and North Dakota ». Plant Disease 91, no 9 (2007): 1105‑8. https://doi.org/10.1094/PDIS-91-9-1105. Khan, J., L. E. del Rio, R. Nelson, V. Rivera-Varas, G. A. Secor, et M. F. R. Khan. « Survival, Dispersal, and Primary Infection Site for Cercospora Beticola in Sugar Beet ». Plant Disease 92, no 5 (2008): 741‑45. https://doi.org/10.1094/PDIS-92-5-0741. Oerke, Erich-Christian, et Ulrike Steiner. « Intra-Leaf Variability of Incubation Period Sheds New Light on the Lifestyle of Cercospora Beticola in Sugar Beets ». Journal of Fungi 11, no 3 (2025): 211. https://doi.org/10.3390/jof11030211. Rangel, Lorena I., Rebecca E. Spanner, Malaika K. Ebert, et al. « Cercospora Beticola : The Intoxicating Lifestyle of the Leaf Spot Pathogen of Sugar Beet ». Molecular Plant Pathology 21, no 8 (2020): 1020‑41. https://doi.org/10.1111/mpp.12962. Rosenzweig, Noah, Linda E. Hanson, Saltanat Mambetova, et al. « Temporal Population Monitoring of Fungicide Sensitivity in Cercospora Beticola from Sugarbeet ( Beta Vulgaris ) in the Upper Great Lakes ». Canadian Journal of Plant Pathology 42, no 4 (2020): 469‑79. https://doi.org/10.1080/07060661.2019.1705914. Rossi, V. Cercospora leaf spot infection and resistance in sugar beet. in Cercospora beticola Sacc. biology, agronomic influence and control measures in sugar beet (2000). |

![]()
Agrément conseil de l’ITB à l’utilisation des produits phytosanitaires n° 7500002.
Assurance RC n° 05421646Y/1025.
Le portail EcophytoPIC recense les techniques alternatives à l’utilisation des produits phytopharmaceutiques.
Organisme agréé Crédit d'impôt Recherche
L'Institut Technique de la Betterave est
membre du réseau Acta
Institut Technique Agricole Qualifié
par le Ministère de l’Agriculture et de l’Alimentation


