

Chaque semaine, les observateurs alimentent le réseau de surveillance des parcelles du Suivi Biologique du Territoire (SBT) pour déterminer la pression sanitaire au cours de la campagne. Il couvre actuellement 214 parcelles dont 105 avec absence de traitement de semence insecticide à base de néonicotinoïdes (NNI), et 109 parcelles avec. Ces données d’observation alimentent l’outil Alerte Pucerons, qui a été développé afin de visualiser en temps réel les seuils d’intervention pour lutter contre les pucerons.
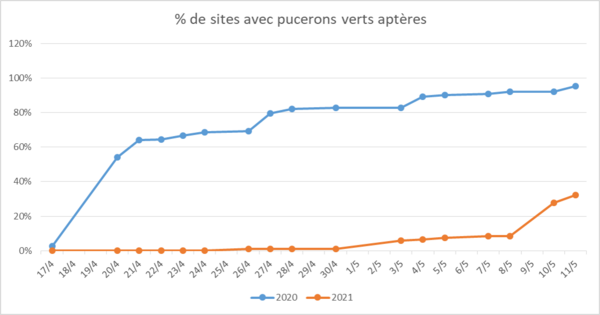
Cette année, les premiers pucerons ont été observés en moyenne à partir du 7 mai. Au 11 mai, ils sont observés dans 48 % des sites sans NNI.
Situation au 11 mai 2021 et comparaison avec les observations arrêtées à la même date de 2020 :
- Comparaison de l'évolution du pourcentage de sites avec pucerons verts aptères
- Détail par sites
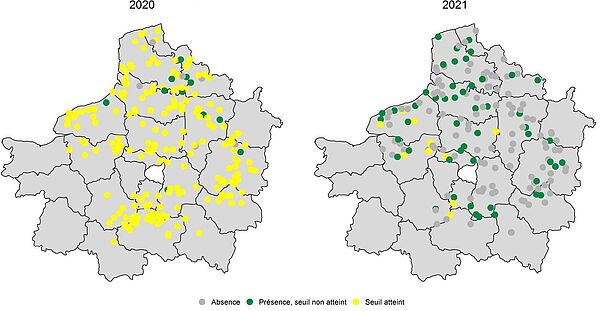
Les illustrations sont réalisées à partir des données issues du BSV et saisies dans Vigicultures©. Le seuil est atteint lorsque 10 % des betteraves sont atteintes par des pucerons verts.
Une différence de pression liée aux conditions climatiques
Les températures météorologiques de janvier et février conditionnent les premiers vols de pucerons verts (Fiche PIC pucerons vecteurs de la jaunisse et jaunisse). L'épisode de gel de février a permis de retarder les premiers vols et de limiter le développement précoce des pucerons verts.
De plus, les températures moyennes basses, inférieures aux normales saisonnières d’avril de cette année ont ralenti leur développement.
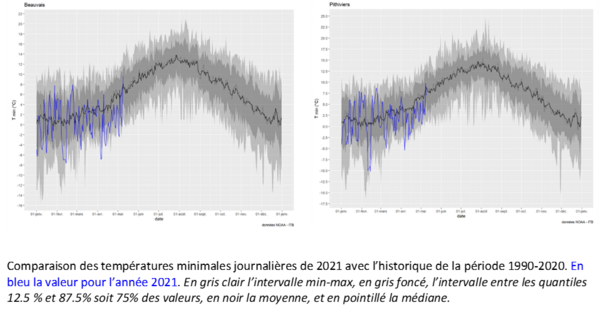
Vague de froid tardive et pucerons
Il n'y a pas de raison de penser que la vague de froid de début avril ait permis de réduire drastiquement les populations de Myzus persicae hivernant sous forme parthénogénétique.
En effet, selon les études la LT 50 (température d’exposition entrainant la mort de la moitié des pucerons) peut varier entre - 6°C et - 16°C (Alford, Blackburn, et Bale 2012; Howling, Bale, et Harrington 1994; Bale, Harrington, et Clough 1988; Clough, Bale, et Harrington 1990) lors d’expositions ponctuelles aux basses températures. D’après les auteurs, cette variabilité peut s’expliquer par différents facteurs qui influencent la résistance des pucerons au froid :
- La sensibilité des pucerons au froid dépend de leur stade de développement, les adultes sont plus sensibles au froid que les larves.
- La sensibilité des pucerons dépend de leur acclimatation au froid. Pour un même clone de pucerons, les individus élevés à 20°C ou 25°C constant ont une sensibilité au froid plus élevée que les pucerons élevés à 10°C constants. Enfin la durée d’exposition aux températures basses a aussi un impact sur la survie des pucerons. Plus l’exposition est longue et moins la LT 50 est basse.
- La mortalité des pucerons est faible lorsqu’ils sont exposés à des alternances de température au cours de la journée. Dans l’étude de Howling (1994), la mortalité de pucerons acclimatés à 10°C constant n’est que de 20 % après 10 jours avec des alternances de températures de 18 h à 5°C et 6 h à - 5°C.
Rémanence des produits néonicotinoïdes en traitements de semences contre les pucerons verts
En 2021, la date moyenne de semis est située au 28 mars. Aujourd'hui après 45 jours de végétation, les betteraves dont les semences ont été traitées avec des néonicotinoïdes sont donc toujours protégées contre les pucerons.
C'est ce que montrent les résultats des essais historiques de l'ITB avec des doses réduites de néonicotinoïdes. Le graphique ci-dessous montre que le nombre de pucerons verts pour 40 betteraves est très faible dans les modalités avec de l’imidaclopride quelle que soit la dose, pendant une durée de 60 à 70 jours après le semis.
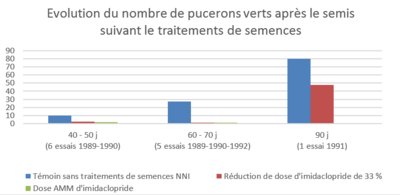
Afin de lutter contre les pucerons en végétation, la dérogation Movento vient d’être acceptée par le ministère. Les informations sont disponibles sur le site internet de l’ITB : une AMM dérogatoire pour le produit Movento
Références bibliographiques
Alford, Lucy, Tim M. Blackburn, et Jeffrey S. Bale. 2012. « Effect of Latitude and Acclimation on the Lethal Temperatures of the Peach-Potato Aphid Myzus Persicae ». Agricultural and Forest Entomology 14 (1): 69‑79. doi.org/10.1111/j.1461-9563.2011.00553.x.
Bale, J. S., R. Harrington, et M. S. Clough. 1988. « Low Temperature Mortality of the Peach-Potato Aphid Myzus Persicae ». Ecological Entomology 13 (2): 121‑29. doi.org/10.1111/j.1365-2311.1988.tb00340.x.
Clough, M. S., J. S. Bale, et R. Harrington. 1990. « Differential Cold Hardiness in Adults and Nymphs of the Peach-Potato Aphid Myzus Persicae ». Annals of Applied Biology 116 (1): 1‑9. doi.org/10.1111/j.1744-7348.1990.tb06581.x.
Howling, G.G., J.S. Bale, et R. Harrington. 1994. « Effects of Extended and Repeated Exposures to Low Temperature on Mortality of the Peach-Potato Aphid Myzus Persicae ». Ecological Entomology 19 (4): 361‑66. doi.org/10.1111/j.1365-2311.1994.tb00253.x.





